
Différence entre la réparation de mésappariement et la réparation d'excision de nucléotide | Mismatch Repair vs Nucleotide Excision Repair
Différence clé - réparation d'incompatibilité vs réparation d'excision de nucléotide
Des dizaines et des milliers de dommages d'ADN se produisent dans la cellule par jour. Il induit des changements dans les processus cellulaires tels que la réplication, la transcription ainsi que la viabilité de la cellule. Dans certains cas, les mutations causées par ces dommages à l'ADN peuvent conduire à des maladies délétères comme les cancers et les syndromes associés au vieillissement (ex: Progeria). Indépendamment de ces dommages, la cellule initie un mécanisme de réparation en cascade hautement organisé appelé réponses de dommages à l'ADN. Plusieurs systèmes de réparation de l'ADN ont été identifiés dans le système cellulaire; ceux-ci sont connus sous le nom de réparation d'excision de base (BER), réparation de mésappariement (MMR), réparation d'excision de nucléotide (NER), réparation de rupture de double brin. La réparation d'excision de nucléotide est un système très polyvalent qui reconnaît les lésions d'ADN de distorsion d'hélice volumineux et les supprime. D'un autre côté, la réparation d'incompatibilité remplace les bases mal incorporées pendant la réplication. La principale différence entre la réparation des mésappariements et la réparation de l'excision des nucléotides est que la réparation de l'excision des nucléotides est utilisée pour éliminer les dimères de pyrimidine formés par irradiation UV et les lésions volumineuses en hélice causées par les adducts chimiques alors que le système de réparation des mésappariements joue un rôle important bases qui ont échappé aux enzymes de réplication (ADN polymérase 1) pendant la réplication. En plus des bases dépareillées, les protéines du système MMR peuvent également réparer les boucles d'insertion / deletion (IDL) qui sont des résultats du glissement de la polymérase pendant la réplication des séquences d'ADN répétitives.
TABLE DES MATIÈRES1. Vue d'ensemble et différence clé
2. Qu'est-ce que la réparation d'incompatibilité
3. Qu'est-ce que la réparation d'excision de nucléotide
4. Comparaison côte à côte - Réparation d'incompatibilité vs réparation d'excision de nucléotide
5. Résumé
Qu'est-ce que la réparation d'excision de nucléotide?
La caractéristique la plus remarquable de la réparation de l'excision des nucléotides est qu'elle répare les dommages aux nucléotides modifiés causés par des distorsions importantes dans la double hélice de l'ADN. Il est observé dans presque tous les organismes qui ont été examinés à jour. Uvr A, Uvr B, Uvr C (excinucléases) Uvr D (une hélicase) sont les enzymes les plus connues impliquées dans le NER qui déclenchent la réparation de l'ADN dans l'organisme modèle Ecoli. Le complexe enzymatique Uvr ABC multi-sous-unités produit les polypeptides Uvr A, Uvr B, Uvr C.Les gènes codés pour les polypeptides mentionnés ci-dessus sont les enzymes uvr A, uvr B, uvr C. uvr A et B reconnaissent collectivement la distorsion induite par les dommages causée à la double hélice d'ADN telle que les dimmers pyrimidiques dues à l'irradiation UV. Uvr A est une enzyme ATPase et c'est une réaction autocatalytique. Puis Uvr A quitte l'ADN alors que le complexe Uvr BC (nucléase active) clive l'ADN des deux côtés des lésions catalysées par l'ATP. Une autre protéine appelée Uvr D codée par le gène uvrD est une enzyme hélicase II déroule l'ADN qui résulte de la libération du segment d'ADN endommagé simple brin. Cela laisse un vide dans l'hélice d'ADN. Après que le segment endommagé a été excisé, un espace de 12-13 nucléotides reste dans le brin d'ADN. Celui-ci est rempli par l'enzyme ADN polymérase I et le surnom est scellé par l'ADN ligase. L'ATP est nécessaire à trois étapes de cette réaction. Le mécanisme NER peut également être identifié chez les mammifères. Chez l'homme, la maladie de la peau appelée Xeroderma pigmentosum est due aux dimères d'ADN provoqués par l'irradiation UV. Les gènes XPA, XPB, XPC, XPD, XPE, XPF et XPG produisent des protéines pour remplacer les lésions de l'ADN. Les protéines des gènes XPA, XPC, XPE, XPF et XPG ont l'activité nucléase. D'autre part, les protéines des gènes XPB et XPD montrent l'activité hélicase qui est analogue à Uvr D dans
E coli.

Qu'est-ce que la réparation d'incompatibilité?
Le système de réparation de mésappariements est initié lors de la synthèse de l'ADN. Même avec la sous-unité fonctionnelle €, l'ADN polymérase III permet l'incorporation d'un mauvais nucléotide pour la synthèse toutes les 10 paires de bases
8 . Les protéines de réparation de mésappariement reconnaissent ce nucléotide, l'excisent et le remplacent par le nucléotide correct responsable du degré final d'exactitude. La méthylation de l'ADN est essentielle pour que les protéines MMR reconnaissent le brin parent à partir du brin nouvellement synthétisé. La méthylation du nucléotide adénine (A) dans un motif GATC d'un brin nouvellement synthétisé est un peu retardée. D'un autre côté, le nucléotide adénine du brin parent dans le motif GATC a déjà méthylé. Les protéines MMR reconnaissent le brin nouvellement synthétisé par cette différence par rapport au brin parent et commencent la réparation de mésappariement dans un brin nouvellement synthétisé avant qu'il ne soit méthylé. Les protéines MMR dirigent leur activité de réparation pour exciser le mauvais nucléotide avant que le brin d'ADN nouvellement répliqué soit méthylé. Les enzymes Mut H, Mut L et Mut S codées par les gènes mut H, mut L, mut S catalysent ces réactions dans Ecoli. La protéine Mut S reconnaît sept des huit paires de bases de mésappariement possibles à l'exception de C: C, et se lie au site de discordance dans l'ADN duplex. Avec les ATP liés, Mut L et Mut S rejoignent le complexe plus tard. Le complexe transporte quelques milliers de paires de bases jusqu'à ce qu'il trouve un motif GATC hémiméthylé. L'activité de nuclease dormante de la protéine Mut H est activée une fois qu'il trouve un motif GATC hémiméthylé. Il coupe le brin d'ADN non méthylé en laissant un nick 5 'au nucléotide G du motif GATC non méthylé (brin d'ADN nouvellement synthétisé).Ensuite, le même brin de l'autre côté de la discordance est entaillé par Mut H. Dans les autres étapes, les actions collectives de Uvr D une protéine hélicase, Mut U, SSB et exonucléase I excisent le nucléotide incorrect dans le monocaténaire ADN. L'espace qui est formé dans l'excision est rempli par l'ADN polymérase III et scellé par ligase. Un système similaire peut être identifié chez les souris et les humains. La mutation de hMLH1, hMSH1 et hMSH2 humains est impliquée dans le cancer du côlon non polypose héréditaire qui dérégule la division cellulaire des cellules du côlon. -> ->
Figure 02: Réparation d'incompatibilité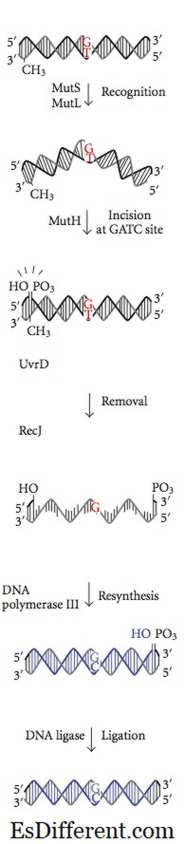
Quelle est la différence entre la réparation d'incompatibilité et la réparation d'excision de nucléotide?
- diff Article Moyen avant Table ->
Réparation d'incompatibilité vs réparation d'excision de nucléotide
Le système de réparation d'incompatibilité se produit pendant la post-réplication. |
|
| Ceci est impliqué dans l'élimination des dimères de pyrimidine en raison de l'irradiation U. V et d'autres lésions de l'ADN en raison d'un adduit chimique. | Enzymes |
| Il est catalysé par Mut S, Mut L, Mut H, Uvr D, SSB et exonucléase I. | |
| Il est catalysé par les enzymes Uvr A, Uvr B, Uvr C, UvrD. | Méthylation |
| Il est essentiel d'initier la réaction. | |
| La méthylation de l'ADN n'est pas nécessaire pour initier la réaction. | Action des enzymes |
| Mut H est une endonucléase. | |
| Uvr B et Uvr C sont des exonucléases. | Occasion |
| Cela se produit spécifiquement pendant la réplication. | |
| Cela se produit en cas d'exposition à U. V ou de mutagènes chimiques, pas pendant la réplication | Conservation |
| Très conservé | |
| Il n'est pas très conservé. | Gap Filling |
| Il est fait par l'ADN polymérase III. | |
| Il est fait par l'ADN polymérase I. | Résumé - réparation d'incompatibilité vs réparation d'excision de nucléotide |
La réparation de mésappariement (ROR) et la réparation d'excision de nucléotide (NER) sont deux mécanismes qui ont lieu dans la cellule afin de rectifier Dommages à l'ADN et distorsions causées par divers agents. Ceux-ci sont collectivement nommés comme mécanismes de réparation de l'ADN. La réparation de l'excision du nucléotide répare les dommages aux nucléotides modifiés, typiquement les dommages significatifs de la double hélice de l'ADN qui se produisent en raison de l'exposition à l'irradiation U. V et aux adduits chimiques. Les protéines de réparation de mésappariement reconnaissent le mauvais nucléotide, l'excisent et le remplacent par un nucléotide correct. Ce processus est responsable du degré final d'exactitude pendant la réplication.
Référence:
1. Cooper, Geoffrey M. "Réparation de l'ADN. "La cellule: une approche moléculaire. 2ème édition. U. S. National Library of Medicine, 01 janv. 1970. Web. 09 mars 2017.
2. "Mécanismes et fonctions de la réparation des défauts d'ADN. "Recherche cellulaire. U. S. Bibliothèque nationale de médecine, n. ré. Web. 09 mars 2017.
Courtoisie d'image:
1. "Nucleotide Excision Repair-journal. pbio. 0040203. g001 "Par Jill O. Fuss, Priscilla K. Cooper - (CC BY 2. 5) via Commons Wikimedia
2. "Ecart de réparation de l'ADN Ecoli" Par Kenji Fukui - (CC BY 4. 0) via Commons Wikimedia
